
《nature》文章:顶级捕食者的恢复减缓了滨海湿地生态系统的地貌衰退
人们认为顶级捕食者的恢复会对植被生态系统及其地貌产生级联效应,但这方面现有的一些证据都只是相关的,而且存在激烈的争论。对此,本研究结合观测和实验数据,揭示了海獭在河口的重新定居产生了营养级联效应,促进了沿岸湿地植物生物量的增加,并抑制了沼泽边缘的侵蚀,从而有助于缓解这一侵蚀过程导致的栖息地和生态系统服务的严重丧失。
几十年来对ElkhornSlough河口的监测表明,尽管该系统的物理压力(即营养负荷、海平面上升和潮汐冲刷)持续增加,但盐沼边缘的侵蚀却随着海獭数量的增加而普遍减缓,表明该系统存在着下行(top-down)控制效应。在五条沼泽小溪中进行的捕食者排除实验表明,海獭会抑制穴居蟹的数量,这种自上而下的效应会增加沼泽边缘的强度并减少沼泽侵蚀。多溪流调查比较了海獭定居前后的沼泽溪流,证实了关键种海獭、穴居蟹和沼泽溪流之间存在相互作用,证明了捕食者对生态系统边缘过程控制的空间普遍性:在海獭重新定居程度较高的溪流中,穴居蟹的密度和边缘侵蚀明显下降。
这些结果表明,营养级衰退可能是造成滨海湿地丧失的一个重要的、但却未得到足够重视的原因,并表明恢复顶级捕食者有助于重建地貌稳定性。
研究背景
地球上几乎每一个生态系统都有顶级食肉动物数量减少的记录。然而,保护行动已经使一些顶级食肉动物得以恢复,包括棕熊、狼和鹰。在恢复过程中,许多食肉动物正在非典型生态系统中定居,而这些生态系统曾被认为远远超出了它们的生存空间(例如,珊瑚礁上的鳄鱼、草原上的山狮和牡蛎礁上的河獭)。在未来几十年中,由于持续的保护努力和气候变化的影响,许多生态系统中大型食肉动物的多样性和丰度可能会出现类似的增长,而气候变化会改变食肉动物的分布。了解和预测捕食者重新进入预期的和意外的生态系统的影响,是生态学和保护工作面临的一大挑战。
在一些顶级食肉动物恢复的例子中,植物生态系统发生了巨大变化。例如,灰狼(Canislupus)和狮子(Pantheraleo)被认为通过对大型食草动物的密度和行为控制,间接控制了它们重新定居的栖息地的植被动态。然而,顶级食肉动物是否是这些生态系统变化的真正驱动力还存在争议。这是因为许多关于顶级食肉动物恢复的研究必然依赖于相关性,而不是综合实验和相关性数据,因此在推断因果关系方面能力有限。以灰狼为例,在美国大黄石地区观察到的植物和河岸结构的变化背后,植物繁殖动态和地貌过程的变化是否与这种顶级食肉动物的恢复有关,这些研究尚待解决。
盐沼生态系统为研究顶级食肉动物恢复的生态后果提供了一个极具吸引力的试验平台,因为盐沼系统既适合进行实验研究,也适合进行观测研究,而且盐沼生态系统目前正在许多地区经历顶级食肉动物的恢复。盐沼对全球温带沿海地区的野生动物和人类也至关重要,它们提供了重要的栖息地、食物、碳储存和海岸线保护。
尽管盐沼具有很高的生态系统服务价值,但它却是地球上最受威胁的生态系统之一,特别容易受到生境改变(如填海造地、水控制结构和海岸线加固)、海平面上升和物种入侵的影响。滨海水域的养分负荷尤其普遍,并已被确认为盐沼消失的一个驱动因素,这是因为养分负荷会影响到沼泽植物,而沼泽植物通过截留沉积物并用根系留住这些物质,自然地抑制了潮汐溪岸(creekbanks,主要潮汐河道的狭窄延伸部分)的侵蚀。具体来说,在富营养化条件下,现场实验表明,盐沼植物会将资源从地下的土壤稳定组织转移到地上的光捕捉叶片和茎上,从而导致溪岸边缘的侵蚀率增加,邻近盐沼的横向损失也相应增加。对全球河口营养负荷和盐沼边缘侵蚀的评估表明,这些过程无处不在,河口营养负荷的增加是全球盐沼广泛、加速和往往永久性丧失(即"地貌衰退")的主要自下而上的驱动因素。
然而,在许多同样的盐沼中,食草动物及其捕食者施加的自上而下的压力也对盐沼的地貌结构和演化产生影响。如果食肉动物不施加控制,高密度的消费者(如螃蟹或蜗牛)会导致盐沼枯死、沼泽边缘侵蚀和下陷。由于过度捕猎和过度捕捞,大多数富营养化河口的食肉动物也明显减少,因此,对植物消耗和/或生物扰动性食草动物释放自上而下的压力可能是造成盐沼溪岸边缘侵蚀的双重机制,从而导致生态系统在人为压力下更加不稳定。
材料与方法
本研究测试了顶极食肉动物(海獭)的恢复对美国加利福尼亚中部ElkhornSlough河口的植物结构和地貌稳定性(即溪岸的横向侵蚀)的影响(图2a)。ElkhornSlough沼泽地是一个以盐沼为主的河口,受到营养负荷、水文变化和海平面上升的威胁。经过数十年的保护,海獭在附近的海带森林中成功恢复,20世纪80年代中期,数个海獭种群在ElkhornSlough重新定居。2000年代,ElkhornSlough沼泽地海獭种群的年度调查显示,一项释放康复海獭的实验性研究计划成功地进一步促进了海獭的恢复,使海獭种群数量增加到几十只。到2000年代末,ElkhornSlough已记录到100多只海獭,并观察到它们在海草和盐沼栖息地觅食(图2b)。
图2研究系统以及海獭数量和溪岸侵蚀的长期趋势。a,ElkhornSlough沼泽地研究地点地图,包括测量溪岸横向侵蚀的地点(黄点,n=196)。b,左图,海獭在一个洞穴严重坍塌的沼泽岸边游泳。右图为ElkhornSlough沼泽中的岸蟹。c,海獭稀少的溪岸,其岸上有大量的滩蟹洞穴。d,沼泽在滩蟹洞穴破坏稳定后坍塌。e,ElkhornSlough的海獭数量(1985-2018年,n=34),显示了海獭向盐沼潮汐小溪扩张的前期、中期和后期。f,潮汐小溪河岸侵蚀时间序列(1937-2018年,n=15)。点代表各个潮汐溪站点。g,潮汐溪平均溪岸侵蚀与ElkhornSlough河槽海獭数量的相位图。e-g中的曲线通过广义加性模型拟合。h,根据贝叶斯模型得出的ElkhornSlough沼泽地海獭数量与潮汐溪流拓宽的预期速率。
虽然ElkhornSlough河口是正在恢复的海獭种群的家园,但由于人类活动,该河口也正经受着巨大的物理化学压力。首先,河口高度富营养化,这对盐沼边缘植物产生了不利影响;富营养化导致植物产生的沉积物稳定根系减少,并助长了能抑制植物生长的藻垫的形成。其次,从1956年到2003年,ElkhornSlough河谷主河道的流速增加了近三分之二。这些高流速使河口从一个间歇性封闭系统变成了永久性开放系统,在1956-2003年同一时期,纳潮量增加了2.5倍,盐沼生境减少了50%。高营养物污染、纳潮量增加和海平面上升的综合影响被认为是造成盐沼边缘侵蚀程度高的三个主要原因。不过,初步观察表明,食物网过程也可能在起作用。
在ElkhornSlough寒冷的河口水域,海獭每天要消耗体重的25%以上才能维持足够的新陈代谢功能。在沼泽潮汐溪流中,海獭主要觅食无脊椎动物,包括岸蟹(图2b)。岸蟹会沿着溪流边缘钻洞(图2c),并食用该地区主要的盐沼植物Salicorniapacifica。与其他盐沼生态系统中的穴居草食蟹类似,岸蟹可能是导致盐沼溪岸不稳定的关键力量(图2d),因为岸蟹的密度相对较高。物理化学胁迫因素(例如富营养化、流速增加、潮汐棱镜增加和海平面上升)和一种标志性顶端食肉动物的恢复(这种食肉动物在河口集中捕食这种丰富的穴居蟹),以及其作为鱼类育苗场和候鸟及留鸟资源的重要意义,使ElkhornSlough成为评估顶端食肉动物恢复对高度受胁迫的沿岸湿地生态学和地貌学影响的一个高度相关的地点。
作者假设,在海獭高密度觅食的潮汐溪流中,海獭会产生营养级联效应,通过抑制沼泽溪岸穴居食草蟹的数量,间接减少河岸侵蚀和盐沼损失。因此,预测海獭的恢复会增强生态系统对人类活动造成的侵蚀力的抵抗力。
对此,本研究采用四种互补的方法来验证其假设:(1)分析在整个河口观察到的海獭数量与盐沼侧向侵蚀之间的十年趋势;(2)在五个潮汐溪流的沼泽边缘进行为期三年的海獭排除实验;(3)对海獭扩张前后的时间序列分析,研究海獭密度、海獭对岸蟹的消耗以及潮汐溪流子集的侧向沼泽边缘侵蚀情况;以及(4)对同一潮汐溪流子集的海獭丰度、岸蟹消耗和侧向沼泽边缘侵蚀情况进行空间分析。
结果与讨论
长期关系
研究首先评估了海獭丰度与同一时期(1985-2018年)潮汐溪流沼泽边缘横向侵蚀之间的时间关系。对历史上潮汐溪流加宽(衡量溪岸侵蚀和增生的指标)的分析表明,1937年至2000年期间,潮汐溪流的加宽率为每年0.09米,之后在2000年代早期(2000-2007年),加宽率增至每年0.35米,之后在2008年至2018年期间又降至每年0.10米(图2f)。溪岸侵蚀的高峰期出现在2000年代初,与1997-1998年厄尔尼诺现象发生后ElkhornSlough和更大的蒙特雷湾地区海獭数量的减少相吻合。此后,在侵蚀减少的同时,2005年至2018年间河口的海獭密度迅速增加(分别为11只和119只)(图2g)。海獭与溪岸侵蚀之间关系的贝叶斯建模结果与海獭抑制溪流拓宽的结果一致,该建模捕捉到了海獭从1992年左右开始在河口的大规模种群扩张。根据模型假设,包括海獭是降低侵蚀率的唯一因素的假设,大约100只海獭与溪岸加宽率从0.38米/年降低到0.10米/年(图1h)有关,每引入一只海獭对溪岸稳定的影响递减。因此,尽管已知驱动ElkhornSlough沼泽地溪岸横向侵蚀的因素(海平面上升、富营养化、潮汐流和潮汐棱镜)持续增加,但沼泽侵蚀率却随着时间的推移不增反减--这种变化与海獭的增加相关,表明对这种地质过程的营养控制是可能的。
捕食者排斥实验
为了进一步评估营养级联假说,并为近几十年来观察到的模式提供机理见解,作者在ElkhornSlough沼泽地的五条潮汐溪进行了为期三年(2013-2016年)的捕食者排除实验(图3a,b)。该实验证实,在沼泽边缘觅食的海獭会产生营养级联,促进沼泽植物地上和地下生物量的增加,并提高土壤容重密度——这些因素已知会影响溪岸的可侵蚀性(图3c-e)。
具体而言,两年后,海獭排除处理区的滩蟹洞穴密度每平方米增加了10只(图3c),而对照区域则没有变化。坑式捕集器中记录的岸蟹数量也对实验处理做出了反应,三年后在海獭排除区观察到的每个捕集器中的岸蟹密度增加了68%。这一趋势在整个实验过程中都是一致的,海獭排除区的岸蟹密度要高于对照样地。与岸蟹密度增加同时发生的是,与对照组相比,海獭排除区的地上植物生物量减少了48%(图3d),地下植物生物量减少了15%(图3e),沉积物体积密度减少了8%(图2b)。
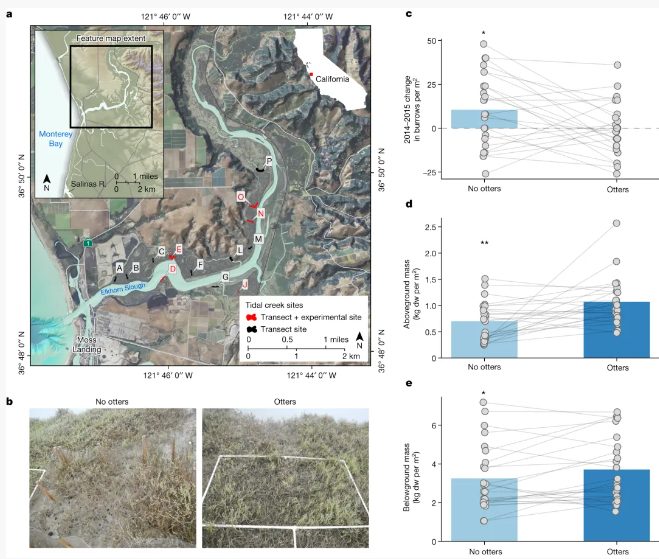

图3实验和观察研究的地点以及捕食者排除实验的结果。a,ElkhornSlough的实验和观察样带图。样带+实验点"表示同时有实验和100米横断面地块的溪流。b,笼养实验的照片,显示笼子将海獭隔开的地块("无海獭";左侧)和海獭可以进入的地块("海獭";右侧)。c-e,为期三年的海獭排除实验结果,响应变量为2014-2015年每平方米岸蟹洞穴的变化(c),以及沼泽地上(d)和地下(e)植物量。
岸蟹的高密度可能通过两种机制加剧沼泽溪岸的侵蚀。首先,通过穴居,岸蟹挖掘出沉积物,而沼泽植物的根茎生长、营养交换和河岸稳定性都依赖于这些沉积物。其次,通过啃食沼泽植物的根部--额外的实验室喂食实验证实了这一过程--岸蟹消除了主要负责控制沼泽溪岸结构完整性的植物性状。尽管地上植被也会受到岸蟹干扰的影响(图3d),但这可能更多是地下物种相互作用的症状。通过实验减少盐沼边缘植物地下生物量的研究表明,这种减少会导致沼泽边缘的侧向侵蚀率增加。
海獭扩张前和扩张后
接下来,研究评估了从重点调查中观察到的海獭丰度、海獭捕食岸蟹的比率以及ElkhornSlough13条潮汐溪的侧岸侵蚀在两个时期内的最新变化:"海獭扩张前"(2009-2012年)和"海獭扩张后"(2015-2017年)。之所以将重点放在这两个时期,因为该系统中的海獭数量增加了106%,从调查期开始到结束,平均数量从62只增加到128只。值得注意的是,2013-2016年的观察期捕捉到了海獭向潮汐溪流和盐沼的大规模扩张。具体而言,在2000年代中期之前,海獭很少使用潮汐溪流觅食,但到2013年,海獭已在溪流中广泛觅食。从海獭扩张前到海獭扩张后,海獭丰度(图4a)和海獭对岸蟹的捕食量(图4b)均有所增加,而溪岸侧向侵蚀率则有所下降(图4c)。结果表明海獭通过溪岸横向侵蚀减少整个河口盐沼损失的假设。
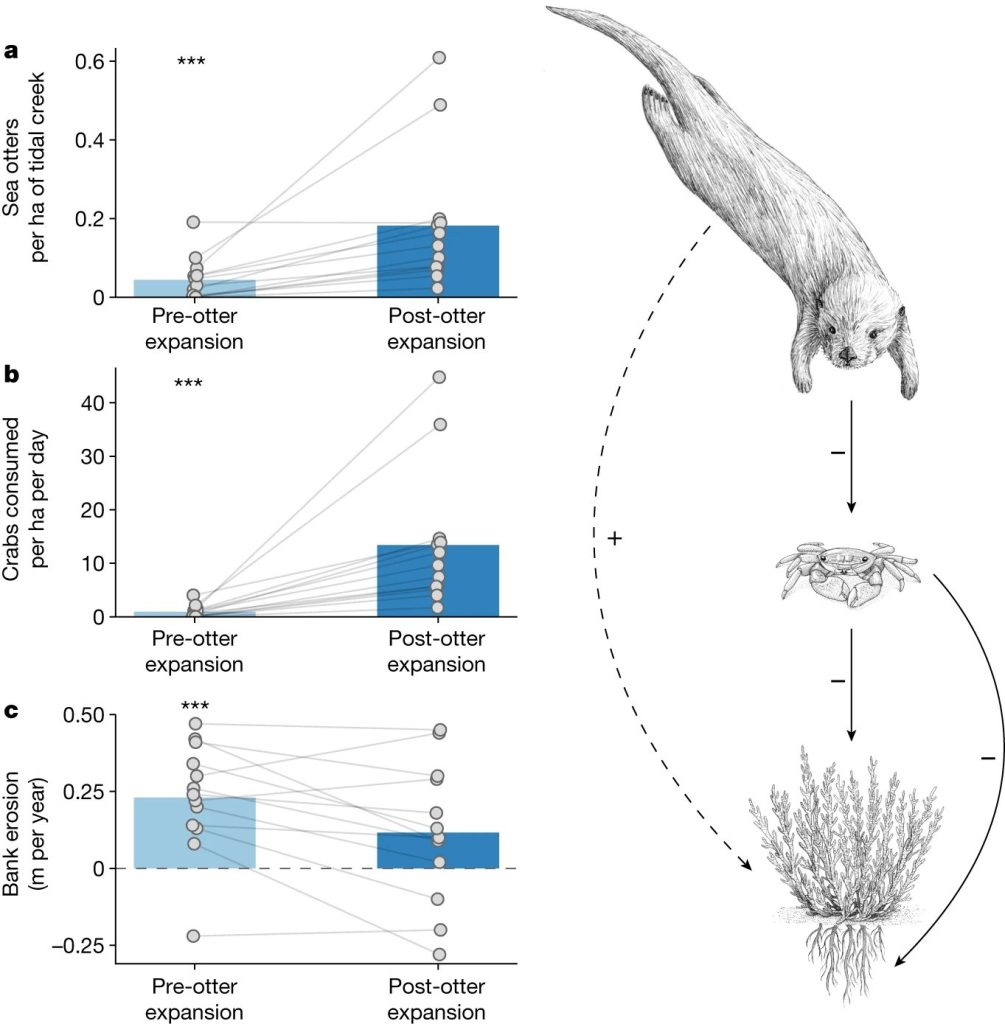
图4潮汐溪流分析结果,比较海獭扩张前后,研究海獭、盐沼生物量和溪岸退缩之间的关系
空间比较
作为对假设的时间评估的补充,本研究还进行了空间比较。具体来说,作者评估了海獭对13条潮汐溪流的利用与岸蟹消耗率(通过海獭觅食调查评估)、岸蟹密度(通过坑式陷阱捕捉岸蟹评估)、植物地下生物量和沼泽溪岸横向侵蚀之间关系的方向、形状和强度。
通过在2013-2015年期间对海獭的使用情况进行目测调查并部署坑式诱捕器,研究发现在海獭密度最高的溪流中,坑式诱捕器捕获的岸蟹数量比海獭密度最低的溪流低约67%。对海獭在潮汐溪流中的觅食行为和猎物消耗的调查证实了这些结果,调查显示海獭的丰度与每条溪流消耗的岸蟹数量呈正相关。海獭每天每公顷消耗多达47只岸蟹(图5a),根据在此期间记录的24,632次觅食潜水计算,约占海獭生物量摄入量的18%。从海獭出没的盐沼区域采集的海獭粪便样本目测结果进一步证实了海獭对岸蟹的摄食:83.6%的粪便样本(n=61)含有岸蟹的身体部分。
空间调查为海獭-盐沼营养级联提供了进一步支持。具体来说,生长在溪岸的盐沼植物的地下生物量随着海獭每天每公顷消耗的岸蟹数量的增加而增加(图5b)。溪岸侵蚀率与海獭每天每公顷消耗的岸蟹数量关系不大;但是,研究观察到一个潜在的异常值。对侵蚀-响应数据的杠杆分析表明,一个离群点对海獭消耗螃蟹与溪岸侵蚀之间的关系有很大影响。从分析中剔除这个离群点后,两者之间的关系仍然显著(图5c)。作者研究了这条离群小溪,发现它不仅有高密度的海獭,而且还有高密度的港海豹。由于海豹在沼泽小溪的河岸上栖息晒太阳,而且它们在我们调查的其他沼泽小溪中并不常见,它们在沼泽河岸上的出现似乎会增加侵蚀率,它们在这条小溪中的高密度是造成离群点的原因。对异常点的这一可能解释,结合统计学上显著的杠杆分析,表明海獭捕食与溪岸侵蚀之间存在空间上的反向关系--这一假设与时间和实验结果一致。
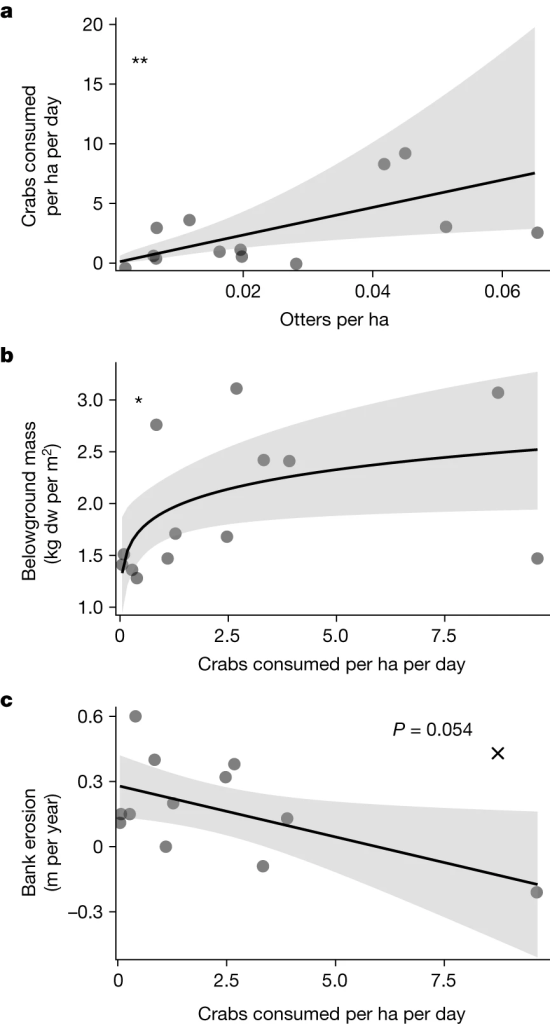
图52013年至2015年ElkhornSlough河口的海獭丰度、岸蟹消耗量和溪岸侵蚀之间的关系。a,海獭丰度的时间趋势(通过每年的航拍调查获得)与海獭每天平均消耗岸蟹的比率之间的关系,该比率通过应用于海獭能量模型的觅食调查进行评估。b,海獭消耗率与地下沼泽生物量(2015年8月测量)之间的关系。c,潮汐溪流的侵蚀与海獭消耗岸蟹之间的关系。标有"x"的点代表一个离群点;这是一个港海豹出没活动频繁的地点。所示的拟合GLM不包括这个数据点。
总结
本研究的时间结果(图2和图4)、空间结果(图5)和实验结果(图3)共同提供了一致的证据,证明自海獭这种顶级捕食者重新定居河口以来,由海獭发起的营养级联已经出现,由此产生的食物网相互作用级联通过抑制穴居、根啃岸蟹减缓了盐沼溪岸的横向侵蚀。
海獭间接控制沼泽溪岸稳定性的证据得到了该系统中三个关键自下而上驱动因素相关数据的进一步支持:富营养化、沉积物负荷和海平面上升。尽管在过去的研究中,这些驱动因素与溪岸边缘的侵蚀呈正相关关系,但研究发现,尽管氮富集、富营养化、沉积物饥饿和海平面上升持续存在并不断加剧,而该系统中溪岸边缘的侵蚀速度已经放缓。尽管2003年是在该系统中测量纳潮量的最近一年,但推测纳潮量在本研究过程中继续增加或保持不变,因为(1)自1947年以来,河口水文变化的遗留问题没有改变;(2)海平面持续上升;以及(3)在本研究过程中测量到ElkhornSlough河槽盐沼的持续侵蚀。
除滨海湿地外,这项工作还对生态学、地貌学和自然保护的理论和应用理解产生了影响。在生态学方面,研究有力地证明,恢复的顶级食肉动物可以改变植物群落、消费者组合和地貌过程,从而使生态系统抵御人为压力。随着其他顶级食肉动物种群的恢复,这一过程可能会变得更加普遍,因为许多自然系统都是与顶级食肉动物共同进化的。在地貌学方面,研究强调了将食物网纳入许多生态系统的地貌演化和对变化条件(如营养负荷和水文变化)的敏感性模型的必要性。从保护的角度来看,研究结果表明,顶级捕食者有可能通过抑制失控的放牧和/或生物扰动效应,缓解沿海湿地,甚至其他系统普遍存在的地貌衰退现象。
这种营养级联的启动可以改变滨海湿地的轨迹,正如物种间的其他积极相互作用(如促进作用)所显示的那样。因此,在以恢复植物生态系统、抑制食草动物加剧的侵蚀和提高生态系统对富营养化的恢复能力为目标时,将捕食者的保护和恢复纳入管理行动可能非常重要。
来源:nature、湿地科学期刊