
《Limnology and oceanography》揭开珊瑚内共体无机碳吸收和利用策略的黑匣子
背景
共生藻科中的甲藻广泛分布,通过光合作用促进珊瑚礁珊瑚的新陈代谢。适应收获和利用光的能力以及 "安全地 "处理光合作用产生的能量是调节其广泛生态成功的关键因素。
Symbiodiniaceae的光合作用是珊瑚礁生态系统的主要能源,而且受到气候变化和环境压力的影响。本篇文章,作者试图探索不同的Symbiodiniaceae如何吸收和利用无机碳(Ci),以及这些策略与其光合适应性和生长策略之间的关系。
研究方法
在培养前,使用浓缩至 100,000 cells mL-1的每个分离物的15 mL等分试样通过测序比对来验证种群培养物的身份。
最初培养了23个不同的分离株,涵盖6个属。除分离物A-2548和E-3420外,所有分离物均已在26°C 的自由活细胞培养条件下保持了至少5年,这两个分离物在实验前进行了低温保存,但在实验前3个月又重新活化并适应了生长条件。
随后选择了三个分离物,分别来自亚热带的共生属Cladocopium和Durusdinium(分别具有热敏感性和热耐受性),以及一个完全来自温带水域的自由生活的分离物(Effrenium 属),以评估生长温度如何调节Ci的吸收和掺入策略。
培养物在250 mL锥形瓶中半连续培养,然后每天对培养物进行细胞计数监测,以确定指数分裂率,并在培养物仍处于指数生长期时取样进行所有后续分析。
用流式细胞仪测量了共生藻的细胞数目、细胞直径和细胞体积,从而计算出细胞的生长率(GR)和细胞大小(CS)。在整个实验过程中定期进行人工细胞计数,以验证自动计数方法的准确性。
碳的吸收与释放
使用放射性标记的碳酸氢钠测定C的吸收。将每种培养物的三份等分样品(10 mL)置于与生长所用的光照强度一致的光照强度下培养,培养时间为60分钟。
培养结束后,将样品置于黑暗中的冰上,以减缓新陈代谢(从而减缓 14C 摄取和细胞掺入),然后用玻璃纤维过滤器轻轻过滤,将滞留在过滤器上的细胞分离到培养基中。添加200μL HCl去除残留的未固定的碳酸氢钠,并使用150μL 25%戊二醛固定样品,然后在通风柜中氮吹。向每个样品中添加10 mL闪烁液,并用力摇晃直至过滤器溶解。然后用液体闪烁分析仪DPM进行计数,从5分钟计数中取平均值。无机碳固定率和有机碳释放率的方法计算如下:


图1计算公式
有机碳量分析
将3 mL的等分试样通过预燃烧的GF/F过滤器,然后立即在液氮中速冻。然后将过滤后放入20 mL硼硅酸盐小瓶中,用200μL HCl酸化以去除残留,然后在60°C真空烘箱中干燥24小时。使用CN分析仪根据EDTA校准标准测定POC,随后将其归一化为细胞密度,分别得到回归后的C、N。POC积累率或NPC通常被视为初级生产率,计算如下:
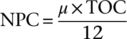
图2计算公式
结果
大多数GPC测量为POC部分,而非DOC部分,因此被细胞保留。共生藻Symbiodinium、Cladocopium和Effrenium分离物的Cexc值普遍高于Breviolum、Durusdinium和Fugacium分离物的Cexc值。有分离物的细胞体积范围很广。虽然已知细胞体积会影响微藻的细胞归一化C吸收率,但本研究中Symbiodiniaceae分离物的细胞归一化C吸收率的变化与细胞体积无关。相反,Cfix或Cexc分别与细胞体积呈负相关和正相关。因此,随着细胞体积的增加,在细胞内保留的碳一般较少,因此表面积与体积的比率降低。
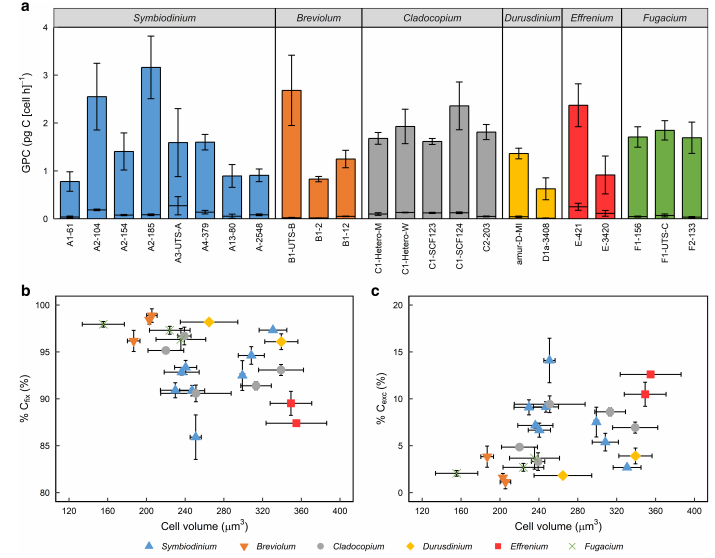
图3在26°条件下,23株共生菌科分离株光合作用1小时后的碳通量(以14C计)。
与GPC相比,NPC表示了细胞对碳的长期吸收和保留,在本研究中各分离物的情况相似。在比较GPC和NPC时,NPC 的碳吸收率始终高GPC。但在整个数据集中,GPC和NPC之间没有显著的相关性,这表明分离物之间GPC到NPC的转化率差异很大。
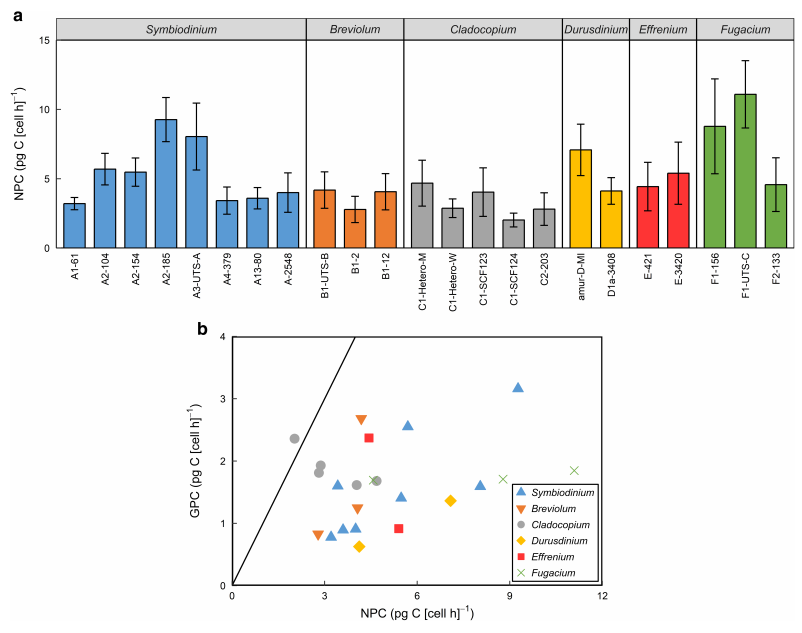
图4 在26°下生长的23株共生菌科分离株的碳产量。
接下来,我们对主要测量碳参数(NPC、细胞体积、Cfix和Cexc)进行了PCA分析,以确定是否可以根据生长、Cfix 和Cexc之间碳吸收的潜在权衡来确定功能组。由此得出,由Cfix和Cexc组成的第一原理成分。在这第一个成分中,Breviolum样本与Effrenium样本被区分开来。第二个成分解释了27.3%的属间差异,主要来自NPC和Cexc,其中Fugacium与Symbiodinium、Cladocopium和 Effrenium区分开来。
次适生长温度对CI同化的影响
三个菌株(C1-SCF124、AMUR-D-MI、E-421)在20℃和30℃以及26℃下生长的Ci分析表明,不同分类群的生长温度表现存在差异。在20-26°C下,C1-SCF124和E-421的生长速度相同,但在30°C下显著降低。相比之下,虽然AMUR-D-MI在20℃时生长完全停止,但在26℃下的生长高于30℃。因此,在26℃时(根据测试的三种温度)生长最好,但通常Amur-D-MI,E-421在较暖、C1-SCF124,E-421在较冷温度下保持生长。生长速度降低的菌株的细胞体积总是更大。C1-SCF124的细胞体积在30℃比26℃大,AMUR-D-MI在20℃比26℃大,E-421在30℃比26°C大。较高的细胞体积对应较低的生长速度,两个变量呈负相关。
进一步比较在20°C、26°C和30°C下生长的三个分离物的GPC与NPC,发现结果与仅在26°C 下观察到的所有分离物的结果截然不同。在30°C生长的所有分离物的GPC值都高于其相应的NPC值,因此GPC:NPC > 1。在20°C和26°C生长的所有其他分离物的比率在0.21至0.66之间。因此,在高温条件下,所有分离物的GPC与NPC密切相关。
最后,PCA分析表明,在所研究的变量(NPC、细胞体积、Cfix和Cexc)中,Cfix和NPC分别更好地解释了第一和第二分量。总体而言,处于最佳生长温度的样本聚集在一起(C1-SCF124 为 26°C,amur-D-MI为30°C,E-421为20°C)。
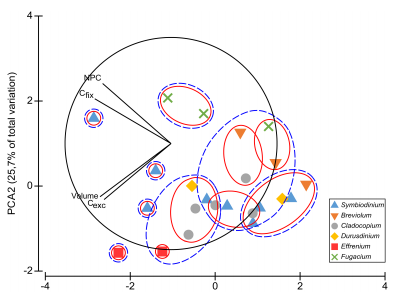
图5 主成分分析
讨论
共生藻的系统发育无法解释C同化的模式
与Symbiodiniaceae系统发育(分类学)多样性的快速增长的知识相反,最终决定这些藻类生态成功的基本生理特性的功能多样性在很大程度上仍未解决。我们使用标准化C摄取和掺入方法对跨越6个属的23个系统发育不同的藻类分离株进行了检查,未观察到任何此类系统发育分组。与光合作用的“暗反应”(即碳反应)相关的性状的这种功能分组的缺乏表明,与光的固定相比这些不同株可能积极地采用不同的生理过程来克服对Ci吸收和利用的进化限制。
与GPC相比,NPC表示了细胞对碳的长期吸收和保留,在本研究中各分离物的情况相似。在比较GPC和NPC时,NPC的碳吸收率始终高于GPC。但在整个数据集中,GPC和NPC之间没有显著的相关性,这表明分离物之间GPC到NPC的转化率差异很大。
在检查的所有生理变量中,只有细胞体积的差异可以解释某些分离株范围内的Ci反应。在我们的研究中,随着细胞体积的增加(即表面积与体积之比的减少),甲藻细胞保留的固定碳较少,释放的固定碳较多。因此,在这种情况下,观察到的更可能代表主动释放,而不是被动溢出。细胞体积未显示出对Cfix的任何影响,这支持了之前在共生藻科中独立于细胞体积的不同C摄取策略的观察结果。
本研究的一个意想不到的结果是,观察到所有分离物的平均NPC约为GPC的三倍(即GPC:NPC <1),这一发现与其他主要浮游植物组的GPC:NPC值报告相矛盾。不过,我们的数值与迄今为止唯一一项对培养的共生藻进行14C代谢的研究一致,该研究报告了确定GPC:NPC比率所需的参数,根据其报告的生长率、POC或14C摄取值,观察到分离物A13-80的GPC:NPC比率为0.36。这一结果耐人寻味,NPC通常被定义为GPC减去暗呼吸、光增强呼吸和光呼吸;因此GPC:NPC值小于1表明存在以下情况之一:
(1)生长所用碳的非光合补充
(2)生长率、POC或14C摄取的方法测定存在偏差
(3)两者兼而有之。
生长环境决定了共生藻的功能多样性
根据C参数(Cfix、Cexc、NPC和细胞体积),在26摄氏度下生长的分离株之间没有观察到明显的功能分组。我们观察到三个关键的共生菌属分离株(C1-SCF124、amur-D-MI 和 E-421)在不同温度下生长时,C获取和生长的功能模式不同,这表明共生菌属中C获取功能多样性的概念可能主要由短期的局部环境驱动,而不是过程的进化分离。我们在次适生长条件下注意到的生理差异似乎反映了不同的固有最适温度,C1-SCF124为 26°C,amur-D-MI为30°C,E-421为 20°C。
只有当分离物的生长温度低于最适温度时,类群间Ci吸收率或C释放率的差异才会明显。这种差异可能表明,细胞对环境干扰的适应主要依赖于上游光合作用途径的调节,如光收集和非光化学淬灭,而不是与下游碳反应相关的途径。即便如此,在温度升高的情况下,共生藻的Cfix并未受损,这与之前的研究结果一致,即藻类的CCM活性主要在次优生长(或 "压力")期间得以维持。然而,Graham 等人(2015 年)研究表明,在热应力作用下,合生藻的Ca活性降低,这可能反映出在某些情况下,Cfix会受到卡尔文循环的影响。因此,为了提高功能群的聚类分辨率,可能需要了解参与C 吸收或光合产物释放途径的其他参数。因此,我们的结果表明,任何功能上的分化都是通过评估它们对光和温度等环境胁迫因素的反应来驱动的。
结论
直接测量C固定量是确定共生藻科碳归宿充分途径,最重要的是可以研究最终可能转移到珊瑚宿主的有机光合产物的释放率。
我们首次发现,在相同的生长条件下,23个Symbiodiniaceae 分离物的C吸收和释放率高度一致,但在次适生长温度下生长时,同一分离物的Ci吸收和释放率却不一致。因此,C "策略"的功能分组主要是由生长环境而非分类多样性驱动的。
因此,共生藻如何应对不断变化的环境可能在很大程度上取决于与光合作用相关的上游过程,如采光和非光化学淬灭的有效性,而不是下游的Ci吸收、同化和释放。
来源:Limnology and oceanography、Academic islander